
Edited by Neil H. Shubin, University of Chicago, Chicago, IL, and approved January 11, 2021 (received for review November 12, 2020)
Author contributions: A.R.M. and S.M.G. designed research; A.R.M., R.E.K., and S.M.G. performed research; A.R.M. analyzed data; and A.R.M., R.E.K., and S.M.G. wrote the paper.
Reconstructions of movement in extinct animals are critical to our understanding of major transformations in vertebrate locomotor evolution. Estimates of joint range of motion (ROM) have long been used to exclude anatomically impossible joint poses from hypothesized gait cycles. Here we demonstrate how comparative ROM data can be harnessed in a different way to better constrain locomotor reconstructions. As a case study, we measured nearly 600,000 poses from the hindlimb joints of the Helmeted Guineafowl and American alligator, which represent an extant phylogenetic bracket for the archosaurian ancestor and its pseudosuchian (crocodilian line) and ornithodiran (bird line) descendants. We then used joint mobility mapping to search for a consistent relationship between full potential joint mobility and the subset of joint poses used during locomotion. We found that walking and running poses are predictably located within full mobility, revealing additional constraints for reconstructions of extinct archosaurs. The inferential framework that we develop here can be expanded to identify ROM-based constraints for other animals and, in turn, will help to unravel the history of vertebrate locomotor evolution.
Our understanding of major transformations in locomotor evolution hinges on our ability to accurately reconstruct how extinct vertebrates walked, ran, swam, and flew. These analyses typically begin with manipulations of fossil limb bones. By evaluating which joint poses are prevented by bony stops or disarticulation, paleobiologists exclude impossible limb configurations from locomotor reconstructions (e.g., ref. 1). Other lines of evidence such as musculoskeletal models, trackway measurements, and robotic simulations can then be used to select which subset of remaining poses to include in reconstructed gait cycles (e.g., refs. 2 and 3).
Ongoing efforts to reconstruct soft tissues in extinct taxa (e.g., ref. 4) and quantify their effects on osteological range of motion (ROM) (e.g., ref. 5) have better equipped paleobiologists to eliminate impossible joint poses from their reconstructions. But might improved ROM estimates also be able to inform which joint poses were actually used, rather than just which ones were not? We propose that the discovery of a consistent relationship between 1) full potential joint mobility and 2) the subset of joint poses used during forward locomotion would allow comparative ROM data to be harnessed in a new way and used to better constrain locomotor reconstructions. Previous studies have suggested that no such relationship exists (e.g., ref. 6), but there have since been substantial advances in the acquisition (7, 8), tracking (9), visualization (7, 10), and quantification (11) of ROM data.
Leveraging these improvements, we designed a case study to identify ROM-based constraints for reconstructions of archosaurian locomotion. Extinct archosaurs such as dinosaurs, pterosaurs, and their kin dominated terrestrial ecosystems for much of the Mesozoic Era. These charismatic “ruling reptiles” have undergone repeated transformations in locomotor posture and mode (12) and are by far the most frequent subjects of locomotor reconstructions (e.g., refs. 3, 13, and 14). Here we carried out a marker-based X-ray reconstruction of moving morphology (XROMM) (15) analysis of unprecedented magnitude to measure nearly 600,000 in and ex vivo poses from the hindlimb joints of two living archosaurs—the Helmeted Guineafowl and American alligator—that represent an extant phylogenetic bracket for the archosaurian ancestor and its pseudosuchian (crocodilian line) and ornithodiran (bird line) descendants. We then created three-dimensional ROM maps (10) to evaluate the relationship between full potential joint mobility and the poses used during locomotion.
Our resulting ROM maps are displayed as Fig. 1. As expected based on previous findings (6, 7), locomotor (here, steady forward walking and slow running) poses comprise only a subset of each joint’s full ROM. However, our improved methodology and comparative approach uncovered three consistent pose usage patterns. First, in all joints studied, pooled locomotor poses are approximately centered along the cosine-corrected flexion−extension (FECC) axis of mobility (see ref. 11), occupying roughly the middle 60 to 75% of the possible range in guineafowl joints, and the middle 50 to 55% in alligator joints. Second, although both animals’ hip joints remain abducted during locomotion, these poses are biased toward the more adducted side of their ROM envelopes, particularly during the weight-bearing stance phase. Finally, all hinge-like joints studied (knees of both species and guineafowl ankle) are biased toward abduction.

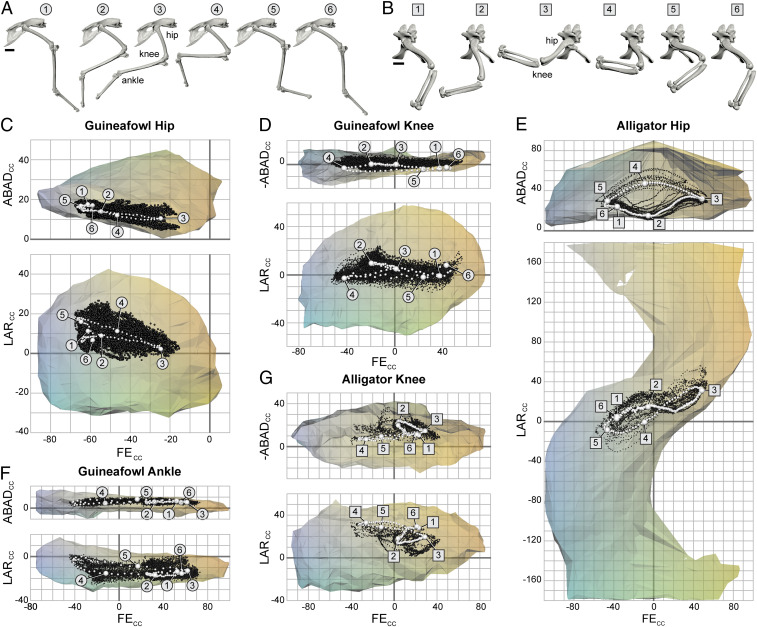
ROM maps comparing full potential joint mobility (polygonal envelopes) to the subset of poses used during walking and running (black points) in archosaur hindlimb joints. A single stride cycle for both the (A) Helmeted Guineafowl and (B) American alligator is highlighted with white points on each ROM map (C–G). A and B depict right hindlimbs in lateral view. (Scale bars, 2 cm.) ROM map axes are FECC (extension toward the right), cosine-corrected abduction−adduction (ABADCC; abduction toward the top), and cosine-corrected long-axis rotation (LARCC; external rotation toward the top). Note that ROM maps for both knee joints plot −ABADCC values to maintain this convention given our joint coordinate systems (see Fig. 2 and SI Appendix). Numbers of poses represented for each joint are (C) guineafowl hip, n = 134,417 (16,324 locomotor); (D) guineafowl knee, n = 127,535 (12,962 locomotor); (E) alligator hip, n = 117,875 (4,499 locomotor); (F) guineafowl ankle, n = 97,695 (12,731 locomotor); and (G) alligator knee, n = 115,897 (4,427 locomotor).
We propose that the pose usage patterns revealed by our comparative analysis of extant archosaur hindlimbs can be translated into constraints for reconstructions of extinct archosaur locomotion. Specifically, we most parsimoniously infer that only the middle 75% of all possible hindlimb FECC excursions should be considered for inclusion in locomotor reconstructions. We also conclude that the pooled set of poses from all reconstructed stride cycles should be centered along the FECC axis of mobility in all joints, and biased toward adduction in hip joints but abduction in hinge-like knees and ankles.
These ROM-based constraints create a substantially narrowed search space within which to reconstruct archosaurian walking and running cycles. The resulting locomotor hypotheses can then be tested using other parameters such as bone strain, energy expenditure, balance, and/or similarity of ground reaction forces to those of extant species (following refs. 1, 13, and 14), ultimately improving our understanding of the locomotion of individual extinct archosaurs and reshaping analyses of archosaurian locomotor evolution (e.g., ref. 12).
Future studies that adopt the inferential framework we establish here will help to determine whether similar constraints can also be applied to other joints (e.g., forelimb joints), taxa (e.g., stem mammals), and modes of locomotion (e.g., flying). When the results of these studies are viewed within a phylogenetic context and integrated with existing evidence from morphology, kinematics, and kinetics, they will inform a broader range of locomotor reconstructions from across the vertebrate tree. We suggest that, in this way, the discovery of additional ROM-based constraints will better illuminate major transformations such as the origins of terrestriality, bipedality, and flight.
Marker-based XROMM analysis was used to measure hindlimb joint poses (simultaneous excursions measured in all three rotational degrees of freedom; Fig. 2) from both in vivo behaviors (15) and intact cadaveric manipulations (8) of the Helmeted Guineafowl (Numida meleagris) and American alligator (Alligator mississippiensis). Procedures conducted with live animals were approved by the Brown University Institutional Animal Care and Use Committee.
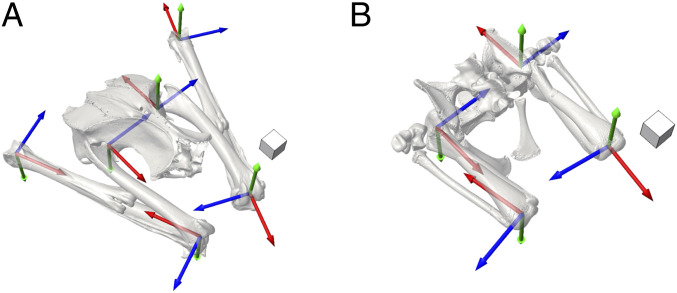
Hindlimb joint coordinate systems for the (A) Helmeted Guineafowl and (B) American alligator. Both species are shown in their reference pose (all joint rotations equal zero). (Scale cubes, 1 cm3.) Rotation about the blue z axis represents flexion−extension, about the green y axis represents abduction−adduction, and about the red x axis represents long-axis rotation. Conventions for positive rotation follow the right-hand rule; note that, whereas abduction is positive at hips and ankles, it is negative at knees.
All measured joint poses were used to create a polygonal ROM envelope representing full potential joint mobility for each joint. Locomotor (steady forward walking and slow running) poses were then plotted within each envelope to create a ROM map (10) in cosine-corrected Euler joint pose space (11). In this space, FECC incorporates information about both flexion−extension and abduction−adduction, resolving the distortion of uncorrected Euler pose space (e.g., refs. 7 and 8) and enabling a quantitative and qualitative assessment of the relationship between joint mobility and the poses used during locomotion.
See SI Appendix for extended materials and methods. All calibration images, X-ray videos, and computed tomography (CT) files are available at http://xmaportal.org/webportal (BROWN20, BROWN58, and BROWN71).
We thank D. Baier for XROMM_MayaTools; J. Hermanson, S. Nesbitt, T. Owerkowicz, K. Roorda, M. Stocker, and R. Wilhite for specimens; J. Lomax, P. Falkingham, T. Roberts, H. Tsai, and M. Turner for assistance with data collection; B.-A. Bhullar, E. Brainerd, and T. Roberts for helpful conversations; and two anonymous reviewers for their constructive reviews. This work was supported by the Bushnell Research and Education Fund, US NSF (Grants IOS-0925077, DBI-0552051, IOS-0840950, DBI-1262156, EAR-1452119, GRFP), Sigma Xi Grant-in-Aid of Research, Society of Vertebrate Paleontology Cohen Award for Student Research, Association of Women Geoscientists/Paleontological Society Winifred Goldring Award, and Brown University Presidential Fellowship.
Calibration images, X-ray videos, and CT files have been deposited in XMAPortal, http://xmaportal.org/webportal (BROWN20, BROWN58, and BROWN71).
1
2
3
4
5
6
7
8
9
10
11
12
13
14
 A new role for joint mobility in reconstructing vertebrate locomotor evolution
A new role for joint mobility in reconstructing vertebrate locomotor evolution
 Facebook
Facebook
 Twitter
Twitter
 Linkedin
Linkedin
 Whatsapp
Whatsapp

