
The authors have declared that no competing interests exist.
The human gut microbiota is a diverse and complex ecosystem that is involved in beneficial physiological functions as well as disease pathogenesis. Blastocystis is a common protistan parasite and is increasingly recognized as an important component of the gut microbiota. The correlations between Blastocystis and other communities of intestinal microbiota have been investigated, and, to a lesser extent, the role of this parasite in maintaining the host immunological homeostasis. Despite recent studies suggesting that Blastocystis decreases the abundance of beneficial bacteria, most reports indicate that Blastocystis is a common component of the healthy gut microbiome. This review covers recent finding on the potential interactions between Blastocystis and the gut microbiota communities and its roles in regulating host immune responses.
Blastocystis belongs to the stramenopile group and is a common single-celled intestinal parasite of humans and a wide range of animals. It is estimated that this parasite has colonized 1 to 2 billion people worldwide, based on epidemiological surveys [1]. Based on analyses of the small subunit (SSU) rRNA gene of Blastocystis, 22 subtypes, which are possibly separate species, have been identified in humans and in a variety of animals [2]. Among them, ST1-9 and 12 have been reported in humans, but ST1-4 are the most common, accounting for more than 90% of human Blastocystis strains [3]. Interestingly, the prevalence of subtypes among regions seems to vary greatly (for a review, see [4]), and different subtypes exhibit remarkable differences in biology, such as drug resistance, immune response, pathogenicity, and effects on microbiota [5–7].
The development and application of next-generation sequencing (NGS) technologies have enabled a better understanding of the role of Blastocystis in the context of the gut microbiome. Recent microbiome studies indicated that this parasite can colonize the human gut for long periods of time without provoking symptoms [8] and that Blastocystis carriers have higher bacterial diversity than non-Blastocystis carriers [9–11], suggesting that it should be considered a commensal rather than a pathogen. In contrast, other studies suggested that Blastocystis is commonly associated with irritable bowel syndrome (IBS) and inflammatory bowel disease (IBD) [12,13]. However, most of these studies have not thoroughly analyzed the etiological role of Blastocystis in the development of IBS or IBD and, in fact, some IBS and IBD patients are not Blastocystis carriers, which in itself should cast doubt on the etiological role of Blastocystis in these syndromes.
The human gut microbiota consists of bacteria, fungi, archaea, and viruses, as well as single-celled eukaryotes [14]. These microbes have a profound impact on human health, metabolism, and the development of the immune system. The majority of human gut studies have focused on the more abundant bacterial components, even though homeostasis in the intestine is maintained through communication and interaction with a variety of microorganisms, such as eukaryotes [15]. Of particular interest is the relationship between single-celled eukaryotes, such as Blastocystis, with the gut microbiota, which has been an emerging research focus in recent years (Table 1).

| Subtype | Method | Gut microbiota composition shifta | References |
|---|---|---|---|
| Blastocystis (ST1-4 and 6) | Metagenomics | INC bacterial richness; Blastocystis mainly found in individuals with Prevotella and Ruminococcus enterotypes | [16] |
| Blastocystis | Real-time PCR | INC in Prevotella and DEC Bacteroides and clostridial cluster XIVa | [17] |
| Blastocystis | Amplicon-based NGS | INC bacterial diversity; INC in Clostridia and Mollicutes (classes), DEC in Bacilli (class); INC in Clostridiales (order), DEC in Lactobacillales (order); INC in Ruminococcaceae and Prevotellaceae (families), DEC in Enterococcaceae, Streptococcaceae, Lactobacillaceae, and Enterobacteriaceae (families) | [18] |
| Blastocystis | Metagenomics | INC in Clostridiales, Firmicutes, and archaeal organisms (Methanobrevibacter smithii); DEC in Bacteroides and Proteobateria | [19] |
| Blastocystis (ST1-8) | Amplicon-based NGS | Not significant | [20] |
| Blastocystis (ST1-4, 8) | Metagenomics | INC Sporolactobacillus and Candidatus carsonella; DEC in Bacteroides | [21] |
| Blastocystis (ST2-3) | Amplicon-based NGS | INC bacterial richness; INC in Prevotella copri, Ruminoccoccus bromii, Debaryomyces hansenii, Mucor mucedo, Aspergillus flavus, Mucor racemosus, and Issatchenkia terricola; DEC in Hymenolepis nana | [22] |
| Blastocystis (ST1-4, 7, 8) | Amplicon-based NGS | INC bacterial diversity and richness; DEC prevalent in Bacteroides enterotyped samples; ST4 was more prevalent in Ruminococcaceae enterotyped samples and associated with Akkermansia; ST3 is inverse | [11] |
| Blastocystis (ST7) | Real-time PCR | DEC in Lactobacillus and Bifidobacterium | [6] |
| Blastocystis (ST1-4) | Real-time PCR | DEC in Faecalibacterium prausnitzii in males; DEC in Bifidobacterium sp. in males with IBS type C | [23] |
| Blastocystis | Real-time PCR | INC in F. prausnitzii/Escherichia coli ratio | [24] |
| Blastocystis | Amplicon-based NGS | INC bacterial diversity; INC in Firmicutes, Elusimicrobia, Lentisphaerae, and Euryarchaeota (phylum); INC in F. prausnitzii and Roseburia sp.; DEC in Actinobacteria, Proteobacteria, unassigned bacteria, and Deinococcus–Thermus | [10] |
| Blastocystis | Amplicon-based NGS | INC bacterial diversity; INC in Clostridiales vadin BB60; DEC in Bacteroidaceae and Escherichia–Shigella | [9] |
| Blastocystis (ST1-4, 7) | Amplicon-based NGS | INC bacterial diversity; INC in Prevotella, Methanobrevibacter, and Ruminococcus; DEC in Bacteroides | [25] |
a Comparison of individuals that were Blastocystis positive with individuals that were Blastocystis negative.
DEC, decrease; INC, increase; NGS, next-generation sequencing.
A retrospective metagenomics approach to studying Blastocystis first revealed that the presence of Blastocystis was positively associated with the Prevotella- or Ruminococcus-driven enterotype and higher bacterial richness, whereas individuals with intestinal microbiota dominated by Bacteroides were much less likely to carry Blastocystis [16]. Moreover, the same research team verified these findings by analyzing another set of fecal samples, observing that individuals who were Blastocystis-colonized alone or along with Dientamoeba fragilis had relatively low abundance of Bacteroides and Clostridial cluster XIVa and high levels of Prevotella [17]. Another large-scale comparative metagenomics study of Blastocystis was recently conducted by Beghini and colleagues, where the presence of Blastocystis was negatively associated with Bacteroides and Proteobateria, whereas strong co-occurrence with Clostridiales, Firmicutes, and archaeal organisms (especially Methanobrevibacter smithii) was observed [19]. The increase of Bacteroides seems to be associated with lower bacterial diversity [26], colorectal cancer [27,28], celiac disease [29], and low-grade inflammation [30], showing that colonization with Blastocystis may be related to a healthy gut microbiota.
Interestingly, recent findings showed that colonization of Blastocystis was strongly associated with increased bacterial richness and various shifts in composition of the gut bacterial microbiota (Table 1). Tito and colleagues showed that the presence of Blastocystis was linked to microbial richness and diversity and found that Blastocystis was less prevalent in Bacteroides enterotyped samples [11]. Similarly, Blastocystis-colonized patients exhibited a higher bacterial diversity and a higher abundance of the Clostridia class, Ruminococcaceae, and Prevotellaceae families, as well as Faecalibacterium and Roseburia genera, while Enterobacteriaceae were enriched in Blastocystis-free patients [18]. Enterobacteriaceae, a family of large gram-negative bacteria, are typically found in a higher abundance in IBD [31]. A similar observation was also recorded by Kodio and colleagues who also observed increase in Faecalibacterium prausnitzii and Roseburia sp. in Blastocystis-colonized children [10]. The Faecalibacterium and Roseburia genera are able to produce butyrate, which is one of the most important metabolites for maintaining colonic health and is the major energy source of colonic epithelial cells [32]. Additionally, butyrate can induce the differentiation of T regulatory (Treg) cells via up-regulation of the Foxp3 gene, to suppress inflammatory and allergic responses [33].
Another study compared the relationship between 3 common intestinal parasites (Blastocystis, Giardia duodenalis, and Entamoeba spp.) and gut microbiota composition in humans. Interestingly, G. duodenalis–positive samples were correlated with a low F. prausnitzii/Escherichia coli ratio, while Blastocystis and Entamoeba spp.–positive individuals related to a higher F. prausnitzii/E. coli ratio [24]. F. prausnitzii is able to reduce the production of the pro-inflammatory cytokines interleukin (IL)-12 and tumor necrosis factor alpha (TNFα) in vitro studies using peripheral blood lymphocytes-derived dendritic cells (DCs) [34]. In addition, Blastocystis was associated with high bacterial diversity and with Clostridiales vadin BB60, while the individuals without Blastocystis had a higher abundance of Bacteroidaceae and Escherichia–Shigella [9]. A previous study reported that high abundance of Escherichia–Shigella appeared to reduce the bacterial diversity and was associated with pro-inflammatory effects [35]. However, other studies revealed no significant bacterial composition differences between Blastocystis-positive and Blastocystis-negative IBS patients [20].
Relationships between Blastocystis and eukaryotic microbiota have also been demonstrated in recent years. Nieves-Ramírez and colleagues [22] evaluated the fecal bacterial and eukaryotic microbiota from 156 asymptomatic adult subjects by amplification and sequencing of the 16S rRNA gene and 18S rRNA genes, respectively, and further compared the composition differences between Blastocystis-colonized and Blastocystis-free individuals. Blastocystis carriers had a higher abundance of Prevotella copri and Ruminococcus bromii, which are commonly found in the human gut microbiome and are often described as either Prevotella-rich or Ruminococcus-rich [36]. Blastocystis colonization was also associated with increases in yeast and fungal species (Debaryomyces hansenii, Mucor mucedo, Aspergillus flavus, Mucor racemosus, and Issatchenkia terricola) and a decrease of Hymenolepis nana [22]. H. nana is a one of the most common cestodes in the human gut and is often associated with pathological consequences [37].
In general, these studies revealed that the presence of Blastocystis was associated with higher gut bacterial diversity and negatively correlated with the levels of Bacteroides. As the higher bacterial diversity is commonly associated with health and lower incidence of inflammatory diseases [38] and the Bacteroides community type has been linked to obesity, inflammation of the lower gastrointestinal tract, and reduced microbiota resilience [39], it suggests that Blastocystis colonization is associated with a healthy gut microbiome.
Apart from these studies that propose a commensal role for Blastocystis, the pathogenic potential of Blastocystis has also been reported (for a review, see [40]). Nourrisson and colleagues suggested that the level of Bifidobacterium sp. was decreased in Blastocystis-colonized male IBS type C patients (IBS patients with constipation based on the Rome III classification [41]) and that healthy Blastocystis-positive individuals had a significant decrease in F. prausnitzii [23]. Similarly, a more recent mouse model study in our laboratory revealed that Blastocystis can decrease the abundance of beneficial bacteria Bifidobacterium and Lactobacillus [6]. Bifidobacterium are known as protective bacteria that have anti-inflammatory properties [42]. Although some strains from Lactobacillus have been linked to sepsis, especially in immunocompromised hosts [43], bacteria belonging to Lactobacillus genus have also been used to prevent opportunistic pathogens infection in the gastrointestinal tract [44]. Blastocystis ST4 colonization in rats can increase the relative abundance of Oscillospira and decrease the level of Clostridium, which is linked to lower amounts of short-chain fatty acids (SCFAs) [45]. In addition, Vega and colleagues showed a significant association between the presence of Blastocystis and Clostridium difficile infection in diarrhea patients [46]. Overall, although these studies reported that the presence of Blastocystis can reduce the abundance of beneficial bacteria, leading to a dysbiotic state, the etiological role of Blastocystis in the development of gastrointestinal diseases, especially IBS, needs further investigation.
There are currently 22 subtypes of Blastocystis, which are widely distributed across human and animals, with some subtypes specific to 1 group but not the other [3] (Table 2). Notably, in linking specific subtypes of Blastocystis to intestinal microbes, only a few subtypes have been well-documented when studying the interactions with gut microbes. For instance, the presence of ST4 in Swedish travelers was associated with higher abundances of the bacterial genera Sporolactobacillus and Candidatus carsonella, while ST3 did not show such significant relationships [21]. Similarly, ST3 and ST4 showed inverse (negative and positive, respectively) correlations to Akkermansia abundance in fecal samples from Flemish gut Flora Project (FgFP) [11], a bacteria that has been linked to delayed onset of obesity and its associated metabolic disorders in murine models [47]. A more recent study showed that ST3 was accompanied by potentially beneficial species, such as Prevotella, Methanobrevibacter, and Ruminococcus [25], whereas co-incubation with lactic acid bacteria has been shown to exhibit strong inhibitory effects on Blastocystis ST3 proliferation in vitro [48]. However, Yason and colleagues showed that presence of ST7 is associated with a decrease of the beneficial bacteria Lactobacillus and Bifidobacterium in a mouse model [6]. Collectively, these findings suggest differential associations between subtypes and host gut microbiota. In light of the enormous inter-genetic variation between different subtypes that exhibited different or even opposite effects [49,50] and only limited studies investigating links between Blastocystis subtypes to gut microbes, future studies should focus on the potential link among Blastocystis and both microbiota community structure and host health, at subtype resolution.
| Subtypes | Hosts | Referencesa |
|---|---|---|
| ST1 | Human; Nonhuman primates; Cattle; Pig; Goat; Sheep; Dog; Cat; Rodents; Birds; Raccoon; Fish; Captive wild animals | [3,51] |
| ST2 | Human; Nonhuman primates; Pig; Dog; Rodents; Birds; Raccoon; Fish; Captive wild animals | [3,51,52] |
| ST3 | Human; Nonhuman primates; Cattle; Pig; Goat; Sheep; Dog; Cat; Rodents; Horse; Raccoon; Captive wild animals | [3,51] |
| ST4 | Human; Nonhuman primates; Cattle; Goat; Dog; Cat; Rodents; Rabbit; Birds; Marsupials; Captive wild animals | [3,51] |
| ST5 | Human; Nonhuman primates; Pig; Cattle; Goat; Sheep; Dog; Rodents; Birds; Captive wild animals | [53] |
| ST6 | Human; Cattle; Pig; Goat; Birds | [54] |
| ST7 | Human; Nonhuman primates; Goat; Dog; Rodents; Birds; Fish | [52,54] |
| ST8 | Human; Nonhuman primates; Marsupials; Dog; Birds; Fish | [51,52] |
| ST9 | Human | [54] |
| ST10 | Nonhuman primates; Cattle; Sheep; Pig; Goat; Dog; Cat; Birds; Marsupials; Fish; Captive wild animals | [52] |
| ST11 | Nonhuman primates; Elephant | [55] |
| ST12 | Human; Cattle; Marsupials; Captive wild animals | [51] |
| ST13 | Nonhuman primates; Marsupials; Captive wild animals | [55–57] |
| ST14 | Cattle; Goat; Sheep; Cat; Captive wild animals | [56,57] |
| ST15 | Nonhuman primates; Sheep; Water Vole | [57] |
| ST16 | Marsupials | [58] |
| ST17 | Cattle; Rodents | [51] |
| ST21 | Waterbuck; Cattle | [56,59] |
| ST23 | Cattle | [59,60] |
| ST24 | Cattle; Sheep; Lama glama | [2,59,60] |
| ST25 | Cattle; Sheep | [2,59,60] |
| ST26 | Cattle; Sheep | [2,59,60] |
a Some references are from reviews because the same subtype has been identified in a wide range of hosts in many studies.
Bold represents the dominant host.
The mechanisms of interaction between eukaryotic parasites and the host have long been the focus of research [61]. Coevolution of gut parasites with the mammalian immune system has promoted the development of complex parasite–host interactomes. In short, these interspecies ties may reveal mutualistic, commensal, or parasitic character alongside many intermediate scenarios, with no explicit cutoff defining the outcome of the host–parasite synergy. Although Blastocystis constitute the most common human-related protist, potentially beneficial or detrimental impacts of these parasites on the host immune system are still under debate [62].
Human-associated Blastocystis represent a genetically diverse component of the gut microbiome [63]. Our previous research demonstrated intra- and inter-subtype variability in terms of ST4 and ST7 pathogenicity [50]. The genetic diversity within the Blastocystis genus is plausibly a critical factor that dictates colonization susceptibility and determines interaction outcomes with the gut immune system. New insights into the Blastocystis–host interactome came from sequencing of the Blastocystis genome. The comparison of ST1-, ST4-, and ST7-derived genomes revealed considerable variation in respect to the genomes’ assembly size, guanine-cytosine (GC) pair content, and the number of protein-coding genes [49]. Proteases constitute an important component of the Blastocystis secretome. Beside engagement in many essential biological processes, proteases are suspected to be potential virulence factors [64]. Importantly, the number and type of protease genes among the subtypes varies greatly [49]. This could provide a possible explanation for the variable clinical significance of Blastocystis.
Most immunological studies on the interactions between Blastocystis and host immune system have focused on ST4 and ST7. The zoonotic subtypes ST4 and ST7 isolates were originally isolated from a Wistar rat and a patient with gastrointestinal symptoms in Singapore, respectively [65,66]. ST4 is the most common subtype in European individuals based on metagenomic studies of the human gut [11,19], while ST7 is rarely found in populations although it was isolated from human. Therefore, we need to note that studies of ST7’s potential pathogenicity may tell us little about how the majority of Blastocystis subtypes present in human gut, for example, ST1 and ST3, and to a lesser extent ST2 and ST4, interact with the human immune system.
Some in vitro studies over the past few decades have been designed to investigate the effects of Blastocystis on host intestinal cells (Fig 1). Cathepsin B, a cysteine protease produced by ST7, has been linked to increased Caco-2 cell monolayer permeability [67]. The trans-epithelial permeability was regulated by tight junctions, which play an essential role in controlling the polarization of epithelial cells and protecting deeper tissues from external microbial pathogen infections [68]. Our previous in vitro study indicated cysteine proteases produced by ST7 induce zonula occludens-1 (ZO-1) and F-actin compromise, in a rho-kinase (ROCK)-dependent manner, in intestinal epithelium [69]. Similarly, ST4 also has the ability to increase the epithelial permeability in IEC-6 cell monolayers [70]. The intestinal permeability of patients with Blastocystis infection was also significantly higher than that of healthy individuals [71]. It is worth noting that the effects of Blastocystis on intestinal cells are mainly based on in vitro studies and may have little bearing on what happens in the human intestine.

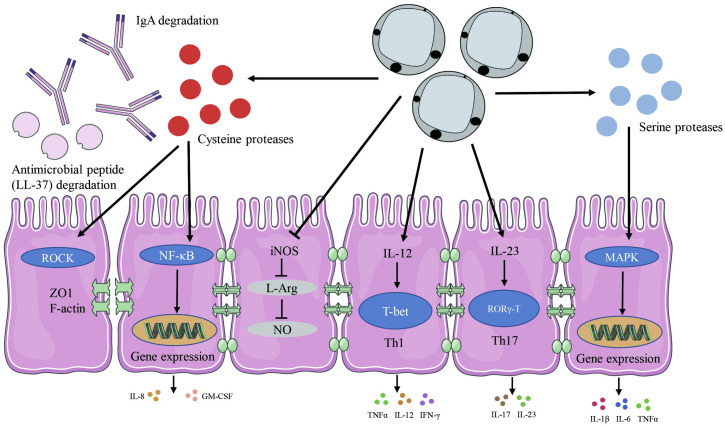
Blastocystis-mediated regulation of immune responses and homeostasis as characterized by studies using in vitro systems and experimental rodent models.
This illustration simplifies the many interactions and pathways involved in Blastocystis colonization or infection in host cells. Cysteine proteases produced by Blastocystis are able to degrade IgA and AMP (LL-37). Blastocystis can also influence the gene expression of pro-inflammatory cytokines by regulating the NF-κB and MAPK pathways. Blastocystis evades host NO antiparasitic response by inhibiting iNOS to convert arginine to NO. Blastocystis can induce Th1 and Th17 cells responses and their signature cytokines release. AMP, antimicrobial peptide; GM-CSF, granulocyte-macrophage colony-stimulating factor; IFNγ, interferon gamma; IgA, immunoglobulin A; IL, interleukin; iNOS, inducible nitric oxide synthase; L-Arg, L-Arginine; MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor-κB; NO, nitric oxide; Th, T helper; TNFα, tumor necrosis factor alpha; ZO1, zonula occludens-1.
The innate immune system constitutes the first line of host defense and plays a crucial role in preventing microbial pathogen infection, while tolerating the normal host flora [72]. Long and colleagues reported that incubation with ST1 modulated the immune response by stimulating the release of the cytokine IL-8 in vitro [73]. IL-8 is a chemokine regulating neutrophil movement as well as, to a lesser extent, other granulocytes [74]. More detailed, mechanistic studies revealed that cysteine proteases produced by ST4-WR1 activate IL-8 gene expression in human colonic epithelial T84 cells in a nuclear factor-κB (NF-κB)-dependent process [75]. Another feature of Blastocystis ST7-B and ST4-WR1-derived cysteine proteases is an ability to degrade immunoglobulin A (IgA), a major immunoglobulin class involved in mucosal defense [76]. Thus, the secretion of cysteine proteases might constitute a parasite’s adaptation mechanism, facilitating colonization by protecting protists in the intestinal milieu against the host’s immune system.
One of the most ancient elements of innate immunity is represented by antimicrobial peptides (AMPs). LL-37 is a 37-amino acid fragment of the human AMP cathelicidin, which exhibits antimicrobial and immunomodulatory activity [77]. In a recent study, employing mouse intestinal explants and the human intestinal epithelial cell line HT-29, it was shown that Blastocystis (ST1, ST4, and ST7) are able to induce intestinal epithelial cells to secrete LL-37 [78]. ST1 and ST4 are susceptible to the effects of LL-37, while ST7 was resistant to the cytotoxic effects of LL-37 through secretion of proteases to degrade LL-37 and an acidified environment to attenuate LL-37 activity [78].
Toll-like receptors (TLRs) are a major family of pattern recognition receptors (PRRs) that play an essential role for protective immunity against infection. TLRs activate downstream signaling cascades which mediate the activation of transcription factors such as NF-κB [79]. In a recent in vitro study, Blastocystis exhibited pleiotropic effects in the modulation of TLR activation by specific ligands (zymosan, lipopolysaccharide (LPS), and flagellin) [80]. Specifically, Blastocystis ST7-B and ST4-WR1 significantly inhibited zymosan-mediated NF-κB activation in human TLR reporter monocytic cell line (THP1-Blue), and neither subtype had any significant effect on flagellin-mediated activation [80]. Interestingly, Blastocystis ST4-WR1 was able to inhibit LPS-mediated NF-κB activation in THP1-Blue cells, while ST7-B was found to augment the effect of LPS-mediated NF-κB activation [80]. However, it should be noted that these results only observed in vitro systems, and future studies should focus on human hosts or other naturally colonized hosts to better understand the role of Blastocystis in host innate immunity.
Blastocystis ST7, but not ST4, induces strong expression of pro-inflammatory cytokines IL-6, IL-1β, and TNFα in murine macrophages mediated by mitogen-activated protein kinases (MAPKs) [7]. The MAPK cascade is one of the most important and evolutionarily conserved signaling pathways and plays an essential role in innate immunity [81]. The MAPKs of mammals can be divided into 3 main families: the extracellular signal-regulated kinases (ERKs), c-jun NH2-terminal kinases (JNKs), and p38 MAP kinases [81]. Blastocystis ST7 triggered ERK and JNK pathways to regulate the expression of pro-inflammatory cytokines in macrophages in vitro, including IL-1β, IL-6, and TNFα [7]. Interestingly, Blastocystis-derived serine proteases increase the pro-inflammatory cytokine expression in a MAPK-dependent manner, while cysteine proteases were able to induce these pro-inflammatory cytokines in a MAPK-independent pathway [7].
Nitric oxide (NO), an endogenously produced molecule, constitutes an important element of the innate intestinal response against luminal pathogens [82]. NO has a strong antimicrobial activity by reacting with a large spectrum of molecules such as DNA, proteins, and lipids [82]. In vitro experiments revealed that ST7-B but not ST4-WR1 is able to inhibit NO production, and this was associated with down-regulation of the transcriptional expression of host cell inducible nitric oxide synthase (iNOS) [83]. Although Blastocystis inhibits an intestinal epithelial NO response in vitro, whether the effect happens in the intact human intestine or in a human intestine damaged by other pathogens (bacterial or viral or parasitic) needs further investigation.
In addition to the impact on innate immune function, recent research also uncovered that Blastocystis plays a role in the adaptive immune system. T helper 1 (Th1) and T helper 2 (Th2) cells and their signature cytokines (e.g., interferon gamma (IFNγ), TNFα, and IL-12 for Th1 and IL-4, IL-5, and IL-13 for Th2 lineage) are important for protection against parasites, intracellular and extracellular pathogens, such as viruses and bacteria [84]. At the same time, potent Th1 or Th2 response at the site of infection may overwhelm the regulatory compartment, which in consequence might lead to chronic inflammation [85]. Yoshikawa and colleagues demonstrated that upon Blastocystis ST4 colonization, rats did not show any pathological lesions; however RNA transcripts for IFNγ, IL-12, and TNFα underwent substantial increases in intestinal tissue of colonized rats [86]. Interestingly, Blastocystis ST4 chronic colonization induces noninflammatory colonic hypersensitivity in rats [45]. Importantly, the colonic levels of IL-6, lipocalin-2 proteins, and GATA-3, a Th2 lineage master regulator, were not changed significantly in Blastocystis-colonized rats [45]. Thus, all these reports indicate a rather controlled engagement of these 2 T helper lineages in an immune response in the context of Blastocystis ST4.
Another more recently identified component of the intestinal T helper compartment comprises CD4 T cells expressing RORγ-T and producing the cytokine IL-17. RORγ-T is a key transcription factor for promoting T cells differentiation into T helper 17 (Th17) cells [87]. IL-17 is known as the signature cytokine of Th17 cells, a CD4 subset contributing to the pathogenesis of various inflammatory diseases [88]. What is important is that the recruitment of CD4 cells toward conventional Th17 phenotype occurs in peripheral organs particularly in the large intestine [89,90]. Furthermore, the site of Th17 lineage polarization does not define a terminal niche for this subset as these cells after differentiation can migrate to the distal locations in respect to the organs of Th17 origin [91,92]. Hashimoto’s thyroiditis (HT) represents an autoimmune disorder with a Th17 lineage playing an essential role in thyroid’s pathogenesis [93,94]. El-Zawawy and colleagues investigated the relationship of Blastocystis and IL-17 in patients with HT. Blastocystis-colonized HT patients had more IL-17 compared to Blastocystis-free HT patients, whereas the amount of IL-17 was significantly decreased after Blastocystis eradication [95]. All these observations correlate with other work in which authors showed increased IL-17 and IL-23 expression in the intestinal mucosa of mice colonized with Blastocystis [96]. Although these 2 studies lack information regarding Blastocystis subtypes, the patterns of cytokine makeup in Blastocystis-colonized HT patients and in intestines of Blastocystis-colonized rodents indicate the potential links between some subtypes and generation of pro-inflammatory Th17 compartment. Hence, functional remodeling of gut microbiomes upon particular Blastocystis subtype colonization or direct interaction of these protists with a host CD4 T immune compartment might lead to the enhanced development and induction of the Th17 pro-inflammatory subset.
In summary, Blastocystis appears to interact with the host immune system at several levels (Fig 1). Although some reports demonstrate that Blastocystis colonization might result in Th1 and Th17 cell responses, the precise mechanism and role of these lineages in controlling colonization-associated pathogenicity has not been clearly explained. The microbiome plays a pivotal role in the shaping and development of the host’s innate and adaptive immune system [97]. Alterations in the composition of the commensal microbiota can influence the frequency of mucosal Treg cells [98]. Indeed, the genus Clostridium, particularly clusters IV and XIVa, can promote accumulation of Tregs, and Clostridium-colonized mice markedly enhanced the differentiation of Foxp3-expressing cells [99]. The immune changes induced by Blastocystis colonization may directly influence the host gut microbiota composition. On the other hand, the remodeling of the intestinal microbiome upon Blastocystis colonization may indirectly modulate the host immune system. Therefore, the complex interactions of the Blastocystis microbiome-immune network requires in-depth studies, which are currently lacking, for mechanistic insights into their roles in gut health and disease.
Based on the above summary of work carried out on Blastocystis, the evidence overwhelmingly points to Blastocystis as a commensal, although a rare subtype (ST7) has shown pathogenicity in in vitro systems and experimental rodent models. Several in vitro and a handful of in vivo rodent experiments reveal that that the common ST4 exhibits mild inflammation and pathology to host tissues. However, we cannot simply regard or designate Blastocystis as a commensal considering numerous genetic variations among subtypes. More microbiome and immunological research should be conducted on humans or other natural hosts at the subtype level to determine if it is a commensal or a pathogen. Furthermore, the mechanisms of the interactions between Blastocystis and gut microbiota are still relatively poorly defined. Since it has been determined that Cryptosporidium infections can affect the metabolite profiling in mouse model [100,101], it would appear appropriate to also ascertain whether Blastocystis colonization is able to affect gut microbial-derived metabolites, such as SCFAs, bile acids (BAs), tryptophan, and/or other metabolites, to maintain the host health and immune homeostasis should be the focus of future research. Importantly, Blastocystis research has mainly used in vitro systems and the experimental rodent models that harbor a divergent microbiota from humans. Therefore, exploring the roles of Blastocystis in impacting immunity requires more mechanistic studies based on human hosts or in other animals with natural colonization.
We would like to thank Dr. Eileen Koh for critical reading and comments on the manuscript.
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
 New insights into the interactions between Blastocystis, the gut microbiota, and host immunity
New insights into the interactions between Blastocystis, the gut microbiota, and host immunity
 Facebook
Facebook
 Twitter
Twitter
 Linkedin
Linkedin
 Whatsapp
Whatsapp

